教育培训在线课程
已有账号,请
如尚未注册?
在线时间:9:00-18:00
010-82782858
澜海源创手机端
随时随地享学习
扫描二维码
关注澜海源创教育公众号
举报
本版积分规则 发表回复 回帖后跳转到最后一页
关注0
粉丝0
帖子148


















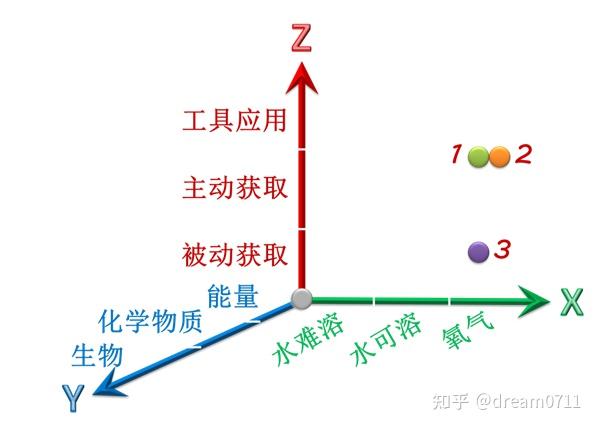 图 1 基于能量供体、能量受体和生物能量传递能力的演变方向和能流矛盾论构建的坐标系
图 1 基于能量供体、能量受体和生物能量传递能力的演变方向和能流矛盾论构建的坐标系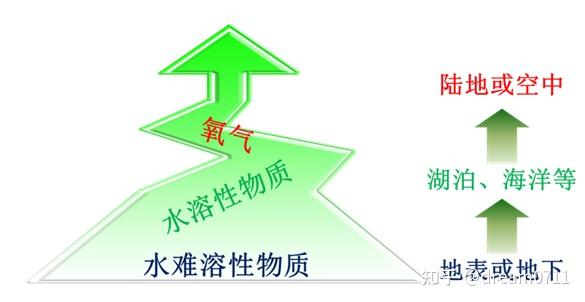 图 2 驱动生物进化的电子受体的演变
图 2 驱动生物进化的电子受体的演变
